Frullania appalachiana R.M. Schuster
Family: Frullaniaceae
Synonyms
none
NatureServe Conservation Status
G1?
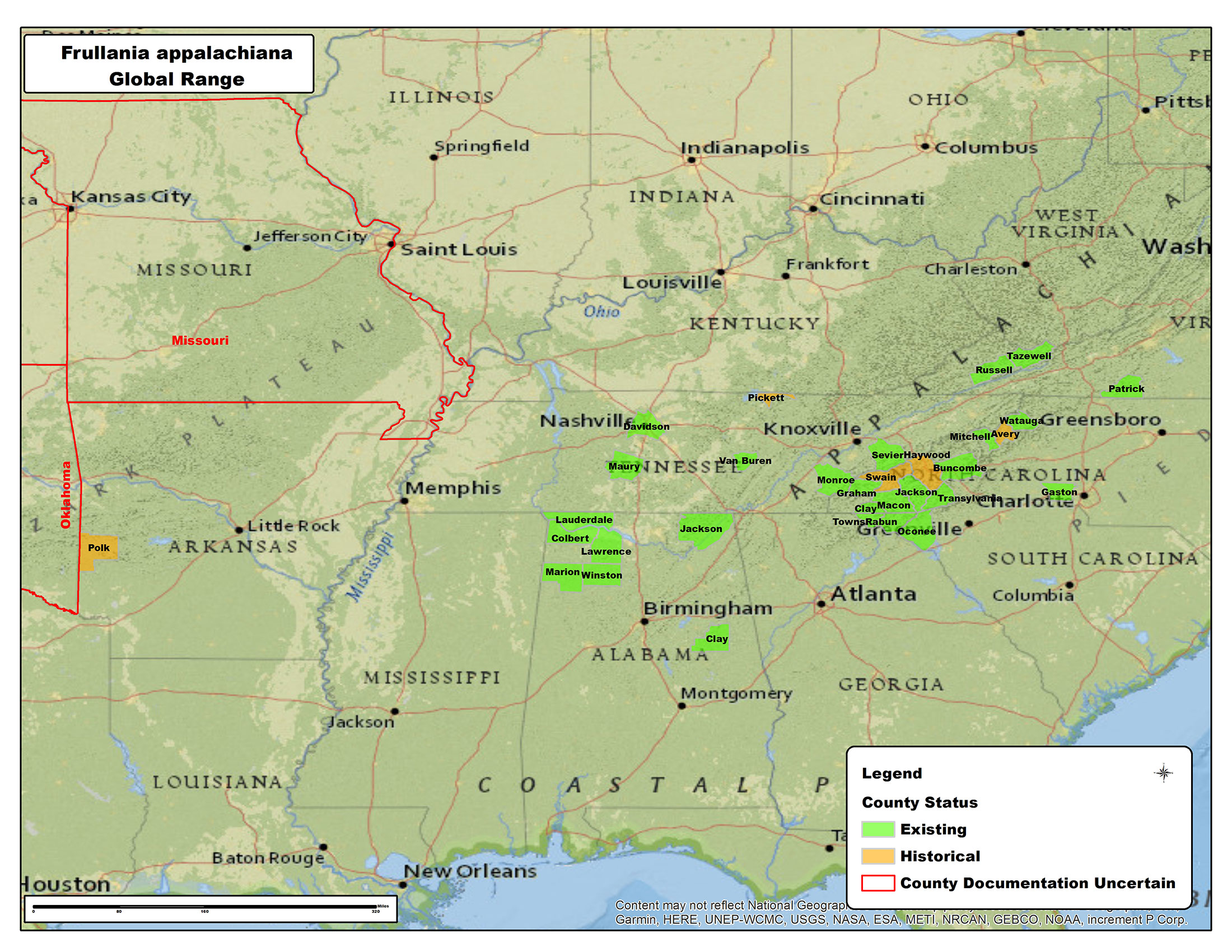
Distribution
Endemic to eastern North America, east of the 100th meridian. Alabama, Arkansas, Georgia, Missouri, North Carolina, Oklahoma, South Carolina, Tennesse, Virginia. (Bryophyteportal.org; Atwood 2017). Originally reported by Schuster (1992) as a localized endemic in the highlands and escarpment gorges of the Southern Appalachians.
Habitat
Frullania appalachiana occurs from low (500 ft) to high (6,000+ ft) elevations in dry oak-hickory forests, ridgetops and upper slopes of northern hardwood forests and Fraser fir-red spruce forests. A key factor appears to be an open understory and conditions that favor periodic low humidity in an otherwise humid climate. The species is not limited to forests and has been found on the trunk of a large willow oak tree lining a city street in Florence, Alabama.
Schuster (1992) noted occurrences on the following trees: Acer spicatum, Betula lutea, Sorbus americana, Castanea dentata, and Liriodendron tulipifera. Additional host trees include the following often found in dryer, open forests: Quercus alba, Q. stellata, Q. falcata, Q. velutina, Q. marilandica, Q. rubra, Q. montana, Carya glabra, C. tomentosa, Acer rubrum, and Ulmus alata. As a rarity found on the following trees, most of which rarely occur in suitable Frullania appalachiana habitat or otherwise presumably have less favorable bark characteristics: Cornus florida, Liquidambar styraciflua, Magnolia acuminata, Aesculus flava, Halesia tetraptera, Fagus grandifolia, Acer saccharum, Nyssa sylvatica, Prunus serotina, Ailanthus altissima, and Quercus phellos.
A few of the closely associated species co-occuring with Frullania appalachiana include F. inflata, F. eboracensis, F. virginica, F. kunzei, and the moss Platygyrium repens. Well formed mats of F. appalachiana visible from a distance are less common than the many small aggregations scattered on bark of oaks, hickories and other trees. On red oak trees with darkly colored bark, small patches of F. appalachiana are easily overlooked, especially when hidden in bark fissures.
Brief Description and Tips for Identification
Shoot width: 0.5-1 mm. Color: brown to olive green. Cell walls in leaf lobe: with intermediate thickenings. Dorsal lobe insertion: cordate. Lobules: about as wide as long.
Dioicous. Perianth with tubercles.
As a member of a complex that includes Frullania eboracensis and F. virginica, F. appalachiana is not always identifiable based on morphology of poorly developed material. In size, F. appalachiana is typical of other bark inhabiting species of Frullania where shoot width is variable, ca. 0.5 mm to nearly 1 mm wide. Appreciably larger shoot widths, averaging nearly 1 mm and slightly larger, characterize F. brittoniae, one of several species sometimes difficult to distinguish. Fortunately oil body number in cells of the leaf lobe easily separate F. appalachiana (oil bodies 2-5) from F. brittoniae (oil bodies 4-8).
The color of F. appalachiana (brown to olive green) has limited value for identification, except to say the color is never extensively grass green as in forms of F. virginica and F. inflata.
Lobules are variable. On good material many lobules will have a closed mouth and some will be large, obscuring half of the dorsal lobe. Underleaves tend to be small, often hardly protruding beyond the flanks of the stem. Schuster (1992) describes the underleaves as “1.6-1.8(2.1) x as wide as stem” yet his figure 796 clearly illustrates underleaves well less than 1.5 x as wide as the stem. Underleaves larger than 1.5x as wide as the stem diameter can be found but on average underleaves are often smaller. The point is, underleaf width varies on a single shoot in most all regional Frullania species. Learning to observe the “tendency” of underleaf width relative to the stem, especially the tendency in the largest of underleaves, should be kept in mind.
The perianth of F. appalachiana is unlike any other regional species due to a combination of features. The dorsal face of the perianth is typically flat (devoid of accessory ridges or plicae) and moderately to heavily adorned with tubercles scattered across the entire face. The beak is short, usually with a non-flaring mouth. The cells of the beak are uniformly quadrate (square) in outline. The female bracts (the last pair of leaves before the terminal perianth) typically have dorsal lobes that remain plane when hydrated.
The perianth of the closely related F. eboracensis is devoid of tubercles. The perianth of F. virginica is distressingly similar. Tubercles scattered across the entire face are common, yet this can vary within a single mat of plants where some perianths are largely devoid of tubercles and others are heavily tuberculate. Fortunately, the perianths almost always have well developed dorsal ridges or plicae. Tubercles may be limited to the ridges or may be heavily produced also occurring on the tissue between the ridges or plicae. The length of the beak of F. virginica varies. Often it is rather long with a flaring mouth. The cells of elongated beaks are elongated-rectangular in outline; however short beaks of F. virginica have similarly quadrate cells as in F. appalachiana. The bracts below the perianth have dorsal lobes that are typically squarrose when wet, although this is not 100 percent consistent for all perianths in a contiguous mat.
Vegetatively F. appalachiana is renowned for its possession of specialized asexual reproduction in the form of caducous leaves, a character shared with F. eboracensis with which it often co-occurs. There is some question as to whether F. virginica occasionally produces rare shoots with caducous leaves, but this appears to be true and is being tested with molecular taxonomy.
Salient Features
- Some lobules with closed mouths
- Lobules of larger, mature shoot sectors large
- Perianths almost always flat dorsally, without supplementary ridges
- Tubercles scattered across both dorsal and ventral faces of the perianth
- Caducous leaves usually present
- Underleaves small
- Dark brown to olive green plants in exposed or open habitats
Perianth bearing material is typically required for confident identification.
References
Atwood, J. J. 2017. Frullaniaceae. Bryophyte Flora of North America, Provisional Publication.
Schuster, R.M. 1992. The Hepaticae and Anthocerotae of North America East of the Hundredth Meridian. Volume V. Field Museum of Natural History, Chicago
Habitat

Frullania appalachiana
Roadside yellow poplars support abundant F. appalachiana and F. eboracensis with smaller amounts of F. brittoniae on these trees at 3800 ft elev. in Buncombe County, North Carolina.

Frullania appalachiana
Various species of Frullania can be found in the dark blotches on light colored tree bark, including F. appalachiana in the above photo.

Frullania appalachiana
F. appalachiana, F. brittoniae, and F. eboracensis occur on this tree. At this scale thay cannot be distinguished.
Habitat

Frullania appalachiana
On trunk of yellow birch

Frullania appalachiana
On trunk of yellow buckeye, Roan Mtn., with Platygyrium repens and Leskella nervosa.

Frullania appalachiana
Trace amounts of F. appalachiana with more abundant F. eboracensis and F. asagrayana on the trunk of a small standing dead tree.
Habitat

Frullania appalachiana
On north facing side of trunk of Quercus phellos, North Wood Ave., Florence Alabama, October 2018. No additional roadside trees in the vicinity were found with this elusive species.
Habit

Frullania appalachiana
Dry specimen with typical color

Frullania appalachiana
The green perianth right of center belongs to F. appalachiana; the many smooth perianths to the left belong to F. eboracensis.

Frullania appalachiana
F. asagrayana running through the center of the photo is larger and shinier than the surrounding F. appalachiana
Habit

Frullania appalachiana
Hydrated specimen in an attempt to show denuded stems due to caducous leaves.

Frullania appalachiana
Hydrated specimen with wiry stems once bearing caducous leaves; a few perianths also visible

Frullania appalachiana
Hydrated specimen, note the female bracts are uncharacteristically squarrose, a feature typical of F. virginia. The perianths are classic for F. appalachiana with a nearly flat dorsal face and scattered tubercles
Habit

Frullania appalachiana
Plants can be very dark and small when growing under highly exposed conditions.
Morphology

Frullania appalachiana
The same perianth shown dorsally and ventrally

Frullania appalachiana
Dorsal side of perianth

Frullania appalachiana
Ventral view of perianth and small branch.
Morphology

Frullania appalachiana
Ventral view of perianth.

Frullania appalachiana
Ventral view

Frullania appalachiana
Ventral view, note the lobules in view are not particularly large
Morphology

Frullania appalachiana
Showing larger lobules

Frullania appalachiana
large lobules with closed mouths
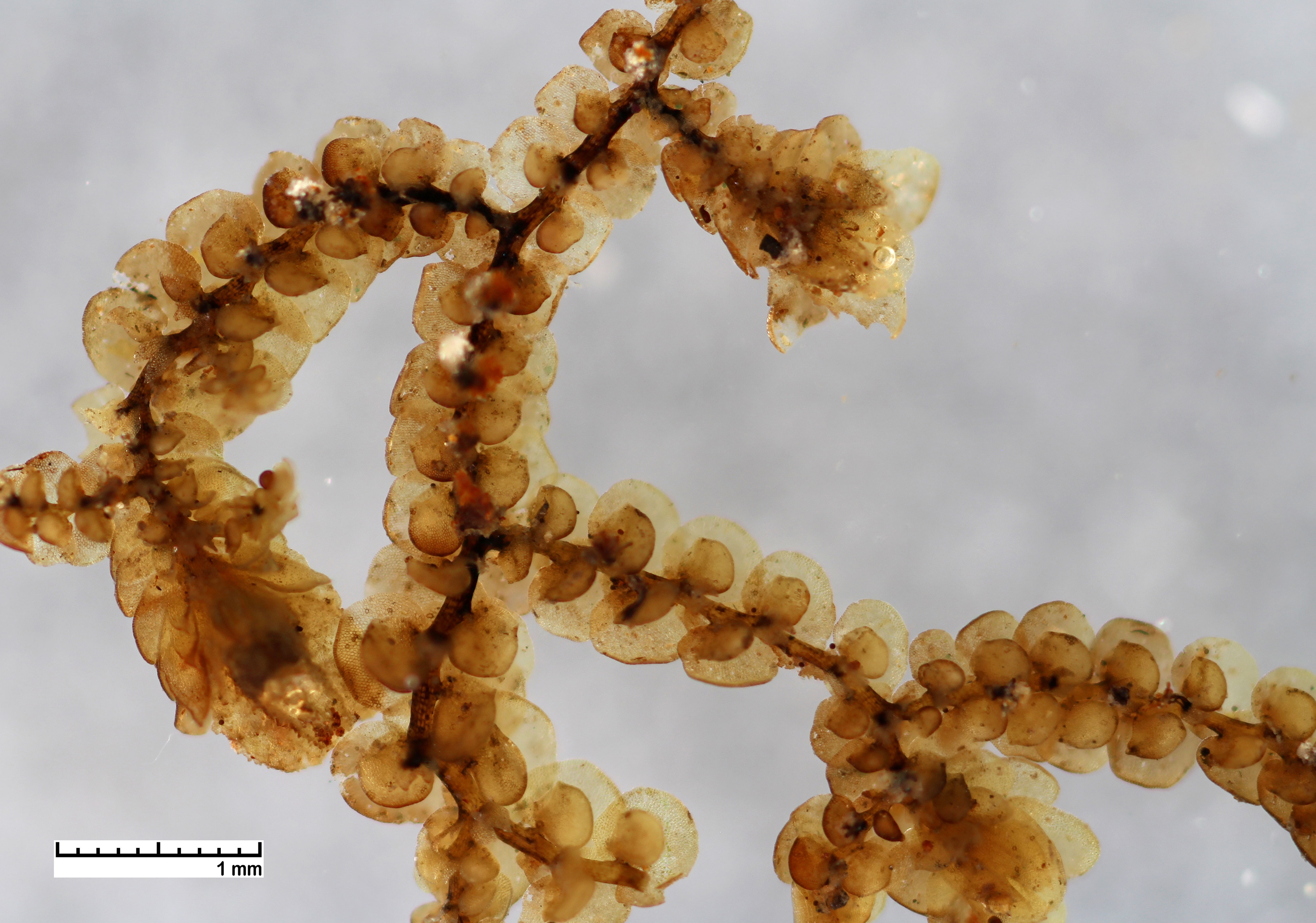
Frullania appalachiana
A 20 year old specimen showing variation in lobule size
Morphology

Frullania appalachiana
Lobules with closed mouths

Frullania appalachiana
Lobules with closed mouths

Frullania appalachiana
Lobules with closed mouths and underleaves somewhat visible
Morphology

Frullania appalachiana

Frullania appalachiana

Frullania appalachiana
Morphology

Frullania appalachiana
Underleaf

Frullania appalachiana
Similar Taxa

Frullania oakesiana
Perianths lack tubercles.

Frullania oakesiana
The species is monoicous.

Frullania oakesiana
The large lobules may lead one to confuse this otherwise very different species with F. appalachiana. F. oakesiana is restricted to the higher elevations in the Southern Appalachians.
Similar Taxa

Frullania brittoniae
Perianths do have tubercles but the beak to the perianth is longer than that in F. appalachiana.

Frullania brittoniae
Underleaves are wider than those of F. appalachiana.

Frullania brittoniae
Oil bodies tend to be more numerous in cells of the lateral leaf lobes, cells with 5-7 oil bodies are frequent.
Similar Taxa

Frullania eboracensis
Perianths lack tubercles.

Frullania eboracensis
Lobules tend NOT to have their mouths strongly compressed (closed).

Frullania eboracensis
Oil body number and wavy cell walls are shared in common with F. appalachiana.
Similar Taxa

Frullania virginica
Perianths can be strongly tuberculate.

Frullania virginica
The pair of female bracts just below the perianth are strongly squarrose when moist.

Frullania virginica
Lobules vary in size but tend NOT to have their mouths strongly compressed (closed).